|
|
とうやく383号(2008年9月号)学術欄HOME > 学術欄 |
|

脳機能発現における はじめに
脳の機能を説明する図やグラフィックアニメーションには多数の神経細胞がそれぞれ神経線維を繰り出して互いに結びあわされて、神経回路を構成している様が描かれ、その中を電気的信号が走って行く様子が示されている。もちろん、それほど詳しいものではないが、それでも他の臓器とはまったく異なった仕組みで機能する脳という組織の複雑さと特殊性を十分に印象付ける「風景画」である。
グリア細胞機能の再発見 ここに登場するのがグリア細胞である。「グリア」というのは神経細胞の間を埋める、セメントとか糊という意味での命名である。日本語としては「神経膠細胞」と訳されている。その存在は細胞としてではなく、膠(にかわ)のような接着剤として認識されていた。しかし、その後、細胞であることが認められ、神経細胞の活動のための環境を整えたり、神経細胞を物理的に支えて形を整えたり、脳に損傷が生ずるとそれを修復するなどの重要な機能をもった細胞として認識されている。しかし、脳機能の本質である情報処理機能に直接関与しているとは考えられなかった。その理由は「グリア細胞は電気的活動をしない」という事実にある。電気的信号の発生は脳機能発現の必須条件であるとすると、グリア細胞が脳における情報処理に直接関わる可能性はまったくないと思われても不思議はない。しかし、最近になってグリア細胞がこれまで考えられてきたように不活発な細胞ではなく情報処理にも積極的に関与することができることが明らかにされ、脳機能発現機構に新しい解釈が迫られるようになってきたのである。
グリア細胞のダイナミックな活動 グリア細胞に情報処理機能があるのではないかと思わせるきっかけを作ったのが、1980年代の初頭に開発された蛍光Ca2+指示薬を用いた細胞内Ca2+計測法の登場である。詳しい説明は省略するが、この試薬を用いれば細胞内のCa2+濃度変化を実時間で計測することができる。筆者は1985年に蛍光顕微鏡と高感度ビデオカメラを組み合わせた装置を独自に開発してCa2+濃度を計測する方法を確立した。この装置を用いて最初に測定したのが、グリア細胞由来の培養クローン細胞の細胞内Ca2+濃度である。セロトニンを適用するとCa2+濃度が見事に上昇したのである。その後、代表的なグリア細胞であるアストログリアがグルタミン酸など様々な神経伝達物質によって顕著なCa2+上昇や振動を生ずることを見出した。我々の発見と前後して、アメリカやヨーロッパでもこの現象は認められた。その結果、グリア細胞は決して静かで、不活性な細胞ではないことが認識されたのである。しかも、グリア細胞の中ではCa2+は波のように動き、別のグリア細胞にも広がってゆくのである。その速度はシナプス伝達に比べて非常に遅い。しかし、間違いなく、脳内にはシナプス伝達とは異なった情報の伝達形式が存在することが明らかになったのである。やがて、アストログリアばかりではなく、オリゴデンドログリアやミクログリアにも同様な応答性があることが発見された。
神経細胞とアストログリアの相互関係
神経伝達物質の遊離はCa2+濃度の上昇を引き金として生じている。もともとアストログリアは成長因子や栄養因子など様々なサイトカインを遊離する能力を持っていることは知られていたので、細胞内Ca2+の上昇はアストログリアが持っている伝達物質様分子の遊離を引き起こす可能性は高いと考えられる。実際にアストログリアがグルタミン酸を遊離することが認められた。これらの発見は神経細胞とアストログリアの間の重要な相互関係の存在を示唆している。すなわち、神経活動に伴って遊離されるグルタミン酸はシナプス周辺に分布するアストログリアを活性化し、その結果上昇したCa2+を引き金にしてアストログリア自身がグルタミン酸を遊離するという促進的な相互関係の存在である(図1)。この仕組みが脳の中に存在するならば、脳機能の発現にアストログリアが積極的に関与している可能性が非常に高くなる。 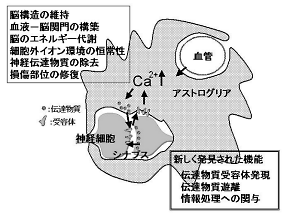
図1 アストログリアは神経細胞と血管の間に存在し、神経細胞の環境の恒常性維持に重要な機能を果たしていることはこれまでもよく知られていた。最近になって、この細胞には様々な伝達物質受容体が発現しており、神経細胞から遊離される伝達物質に反応して細胞内Ca2+上昇させること、さらに自らが持つ伝達物質(グルタミン酸、ATP、D-セリン)を遊離して、シナプスにおける情報処理機能に積極的に影響を与えることが明らかにされてきた。
トライパータイトシナプス
これらの事実は脳の機能が神経細胞で構成される回路のみから発現すると考えることの問題点を指摘している。すなわち、アストログリアがシナプス活動に応じて刺激され、その結果、遊離される伝達物質によって神経細胞の活動をコントロールするという図式が成り立つのである。面白いことにアストログリアはグルタミン酸の他にATPを大量に遊離する。このATPはエネルギー物質としてではなく、情報伝達物質としてシナプス活動を強く抑制することが知られている。アストログリアの形態と存在の様式を考えると、このATPが脳機能全体の興奮性を包括的に制御している可能性が考えられる。極めて複雑で膨大なシナプス構造から成り立つ我々の脳機能が暴走することなく、安定に機能できるのはこのような包括的な制御機構によるのかもしれない。しかし、どのような場面でグルタミン酸とATPによる、促進的調節と抑制的調節が機能しているのかという疑問にはまだ明瞭な解答は与えられていない。
おわりに 本稿では紙幅の制限から十分述べられなかったが、神経細胞は直接血管と接触することはなく、この間にはアストログリアが介在している。脳機能が高まるとそれを感知して、血管を拡張させる機能も持っている。現在は、神経細胞とグリア細胞と血管とを同時に考慮しながら脳機能を考えるべきだと主張している研究者も多い。また、アストログリアには多様な形や機能がある。個体の発生の段階から、病態までを考えると、高次脳機能をニューロンとそれが作る回路網のみで語ろうとしてきた脳研究はまだまだ骨格の部分に過ぎないことを痛感させられる。また、本稿ではアストログリアのみを取り上げたが、その他のグリア細胞、ミクログリアやオリゴデンドログリアなども重要な機能を持っており、脳機能発現への寄与が発見されている。これからのグリア研究の成果が、「脳の風景画」の中に細やかに書き込まれて行くことになる。一筆ずつ新しい要素が書き込まれる度に新しい輝きをましてくる「脳の風景画」は例えようのない魅力を秘めた荘厳なものであるに違いない。それが何年後に完成するのか予測もできないが、是非、見てみたいものだと考えている。
|
|